 | La Spongieuse | Vues 5450 fois |
|
||||
|
|
||||
|
||||
|



|
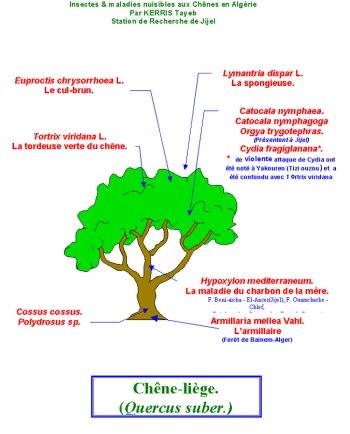
Insectes et maladies nuisibles aux chênes en Algérie
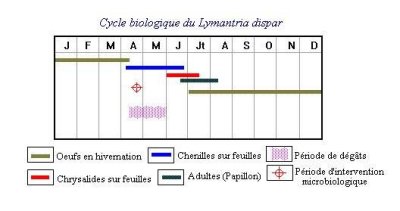
Biogéographie du Lymantria dispar Originaire de l’Extrême-Orient (Japon, Corée), le Lymantria dispar (Lépidoptère Lymantriidae) s’est propagé progressivement vers le Nord de la Chine, la Mongolie, la Sibérie, le Turkestan, le Caucase, le Moyen-Orient; l’Europe, et l’Afrique du Nord (BESS, 1961 et FRAVAL et al., 1989) Dans la région méditerranéenne, l’insecte se retrouve essentiellement sur le chêne-liège et le chêne-vert. Au Nord de l’Europe et aux États-Unis, elle peut se développer aussi bien sur résineux que feuillues, soit sur plus de 458 espèces végétales (FORBUSCH et FERNALD, 1986). En Yougoslavie, le Lymantria dispar vit sur plus de 208 espèces végétales (JANCIVIC, 1958) Le Lymantria dispar possède deux moyens pour assurer sa dispersion spatiale. Par voie éolienne, les jeunes chenilles (L1) légères et couvertes de longs poils qui augmentent leur surface pour parcourir de grandes distances et infester des massifs forestiers. Elles peuvent se déplacer jusqu’à 30 km par jour (JOBIN, 1979). Plus la dispersion est importante, plus la population est en bonne santé (CAPINERA et BARBOSA, 1976). La seconde voie de dissémination de l’insecte, sous forme de larves ou oeufs, se fait par le transport des divers substrats comme le dessous des véhicules, les sacs des touristes etc... (JOBIN, 1979).
Le Lymantria dispar qui est un insecte polyphage
s’alimente sur plusieurs essences (clic |







|
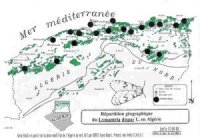
La distribution spatiale de Lymantria dispar en Algérie couvre l’ensemble des écosystèmes à chêne-liège et à chêne-vert. (voir carte de distribution géographique du Lymantria dispar en Algérie). |
|
|
Historique des épidémies antérieures
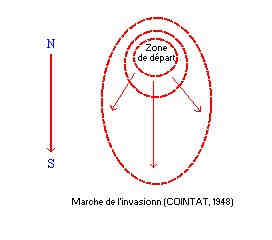
La première infestation signalée est celle qui a sévi dans la forêt d’Edough -Annaba, entre 1923 et 1926. Une seconde infestation de l’insecte fût observé à M-sila en 1925 (BALACHOWSKY, 1935). Une infestation a ensuite été notée en 1934, dans la chênaie verte de Tlemcen et, enfin en 1961, au Sud-ouest d’Azazga (KHOUS, 1993). En Algérie, les attaques de Lymantria dispar sont essentiellement concentrées dans les wilayas suivantes, les taux d’infestation ont été évalués en 1978 (HAMRA-KROUA), et en 1986-1987 (HAMAMI, 1987): Dégâts Pendant les phases de gradations, les chenilles occasionnent des dégâts considérables qui conduisent à la défoliation complète des arbres. On peut comparés l’invasion de la spongieuse à un grand incendie, la marche est à peu près la même et le vent a une grande influence en balançant les chenilles au bout de leur fil d’un arbre à un autre. L’invasion passée, la forêt à un aspect d’une forêt incendié. Les colonies de chenilles, suivent les besoins de leur nourriture, se déplacent et rayonnent autour du foyer (zone de départ). Par ailleurs, pour certaines espèces, on note des perturbations plus fortes, les chênes voient leur glandée compromise. Quant au chêne-liège, une forte attaque fait obstacle au démasclage. Au U.S.A Une défoliation à 100 %. A fait une perte de la croissance radiale de 40 % et de 50 %. En Roumanie une mortalité de 50 % (KULMAN, 1971); Et en Yougoslavie une forte attaque sur chêne pédonculé de 80 ans, suivi d’une deuxième défoliation résulte une perte de croissance de l’ordre de 30 %. Ce déficit de production se maintien ou même s’accentue l’année suivant le sinistre (KLEPAC, 1959). Importance économique de Lymantria dispar Sur le chêne-liège en Europe, une défoliation de 50 % et à 100 % provoquerait respectivement une diminution de l’accroissement en hauteur de 19 % et 63 % la première année et, de 49 % et 91 % la seconde (CAMBINI, 1972). La défoliation se répercute aussi sur la production du liège qui diminue de 42 % et 60 % la première année et, de 10 % et 32 % la seconde année (CAMBINI, 1972). Au Canada d'après JOBIN, (1979) souligne que deux ou trois pullulations pourraient entraîner la mortalité des chênes et qu’une seule défoliation y achèverait un sujet déficient. En juin, en s’attaquant aux fleurs femelles du chêne liège, la spongieuse compromet la fécondation et par conséquent les glandées et la régénération de l’espèce (ZERAIA L., 1988). L’action de Lymantria dispar peut affaiblir d’avantage les peuplements et faciliter l’installation des insectes xylophages et des champignons lignivores. Les défoliations de Lymantria dispar entravent les opérations de démasclage, les sujets défoliés, déjà affaiblis, ne doivent pas faire l’objet de déliègeage. |
|



|
|
||
|
éclosion des oeufs |
||
|







|
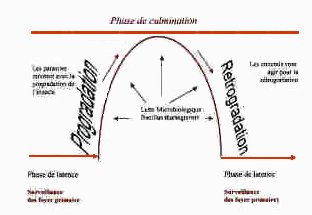
Les ennemis naturels de Lymantria dispar Les Invasions apparaissent soudainement, sévissent pendant 1 à 3 ans puis disparaissent brusquement, se sont les biens fait des ennemis naturels du Lymantria dispar. |
|||||||||||
|









|
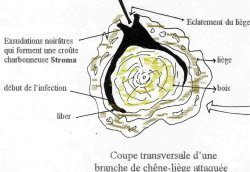
La polyédrose du Bombyx disparate (Maladie virale) a été découverte en 1907 dans les foyers de la Nouvelle Angleterre ou les chenilles ravageaient déjà des milliers d’hectares. Elle a été observé en abondance dans la forêt de l’Edough 5Algérie) en 1925, et elle se déclencha brusquement dans les colonies a leur maximum de densité d’invasion (BALACHOWSKY, 1951). La maladie se transmis par la gestion de nourriture contaminée , aussi les parasites et prédateurs jouent un rôle de transmissions ALLEN (In BALACHOWSKY, 1951), a observé que les mouches sarcophagidae étaient attirées par les chenilles malades et avaient tendance, de cette manière à provoquer la maladie au milieu des colonies saines. La maladie se transmit également à la descendance par les œufs des papillons femelles contaminées qui constituent ainsi une réserve de virus. (BALACHOWSKY, 1951). D’après BERGOLD (In BALACHOWSKY, 1925), qu’il est probable que dans les années à venir une méthode de propager pratique permettra de propager plus efficacement cette maladie et de détruire les invasions de chenilles dès la première année d’invasion.
|
||
|
Stratégie de lutte et recommandations Il existe des forêts dite « sensibles aux attaques des chenilles », qui servent de base de départ à l’invasion, ils sont toujours attaquées les premières, et qui souvent sont les seules attaquées si des solutions sont proposées et misent en pratiques, il devront avant tous être porter sur ces forêts dite « sensible ». Comme on l’a dit les colonies de chenilles suivent les besoins de leur nourriture, il se déplacent et rayonnent autour de la zone de départ du foyer qui se trouve d’après COINTAT dans les vieux taillis. DoncLa localisation et la délimitation des foyers primaires d’infestation du bombyx disparate demeurent une condition sin qua non si l’on veut réprimer ses infestation Alors, Il est fortementrecommandé en toute urgence de localiser les foyers potentiels du bombyx disparate à l’échelle Nationale sur cartes de distribution, et ce avant la future infestations. Cette procédure permettra en effet de mettre en place un réseau d’alerte et de suivi, de prédire les gradations de l’insecte si on souhaite programmer suffisamment à l’avance des interventions de lutte. Par contre nous déconseillons l’intervention sur les massifs envahis, les populations migrantes étant de toute manière condamnées à s’écrouler. Au niveau des foyers en rencontre l'ensemble du cortège parasitaire, étant donné la position favorable du point de vue écologique dans ces foyers (préférendum écologique), équilibre entre l'insecte et son hôte (arbre). La surveillance des foyers d'infestation est indispensable afin de mettre en place un modèle prédictif, et de mieux géré l'exploitation du liège. Si un massif sera attaqué, ne pas exploiter la parcelle, étant donné que les arbres seront en état de faiblesse. Par contre nous déconseillons l’intervention sur les massifs envahis, les populations migrantes étant de toute manière condamnées à s’écrouler. |

|
||||||
|
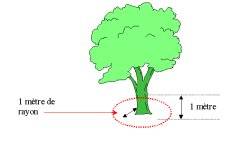
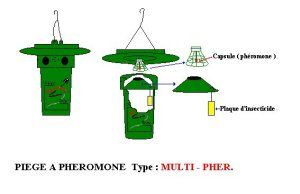
Avertissement Procédures et méthodes d’échantillonnage Détection précoce des chenilles La détection précoce du Lymantria dispar au stade larvaire vise à établir le plus grand nombre possible de points d’échantillonnage sur le territoire, afin de ne retenir que ceux dont la présence de l’insecte est jugée significative. La détection précoce est plus aisée dès les premiers âges larvaires (début Avril - juste après le débourrement des feuilles du chênes), car elles sont libres sur le feuillage durant le jour; plus tard (fin-juin - Disparition des chenilles sur le feuillages, on observe quelques rares individus ici et là sur les troncs d’arbres), elles ont tendance à se regrouper sur les branches et le tronc durant le jour pour se nourrir seulement la nuit (sauf en cas de forte densité, elles se nourrissent jour et nuit.). · Matériel : sécateur, drap d’échantillonnage. · Unité d’échantillonnage : 03 extrémités de branche de 01 mètre par arbre. · Seuil de tolérance (T) : 15 chenilles : Niveau 0 X = 0 négatif Niveau 1 X < 15 présence non significative Niveau 2 X ³ 15 présence significative. Alerte · Nombre d’arbres observés : 100 · Nombre d’arbres échantillonnés : 10 Pontes Le niveau de population de la spongieuse gouverne son comportement de ponte. La femelle dépose ses oeufs sur le sol forestier uniquement en cas de fortes densités de population. Délimitation des parcelles : Elles s'évaluent d'année en année par le dénombrement de masses d’œufs, entités visibles d'Août à Avril. La présence des pontes sur les branches ou les troncs est le critère de reconnaissance le plus typique de Lymantria dispar. La longueur de la ponte indique le niveau de la population puisqu’elle est directement proportionnelle à sa fécondité. Donc le nombre et la taille sont un bon indicateur de la gradation; elles sont plus petites et de relief moins accentué en période de régression. La hauteur des pontes dans l'arbre est en fonction de la densité du couvert végétale. En peuplement dense, les pontes sont hautes, alors qu'en peuplement ouvert, elles se situent en bas de l'arbre . La densité des pontes est plus élevée quand l’éclairement est plus grand, donc aux lisières des peuplements et en bordures des chemins et des clairières. Éviter la prise de données en bordure. D’après KHOUS (1993), les masses d’œufs pouvant être divisées en 3 catégories de tailles : Catégorie I 0 à 20 mm renferment des effectifs d’œufs de 0 à 180 oeufs Catégorie II 20 à 30 mm renferment des effectifs d’œufs de 180 à 280 œufs. Catégorie III 30 à 40 mm renferment des effectifs de 280 à 360 œufs. |


|
||
|
A l’aide d’un pied à coulisse mesuré le diamètre maximum de chaque ponte jusqu’à concurrence de 60 pontes consécutives (en raison de la difficultés de retrouver 60 pontes en se contente du maximum de ponte que vous trouvez, peu importe l’essence hôte). Sur les fines branches de moins de 2 cm de diamètre, les pontes sont plus allongées et étroites, elles doivent ignorés lors des mesures. · Matériel : pied à coulisse de 0,05 mm. · Unité d’échantillonnage : station · Nombre de pontes dans la station d’observation : 60 pontes Il est important que chaque observateur puisse reconnaître facilement l’insecte sur le terrain. Les pontes localisées sur les fines branches sont plus allongées et plus étroites, elles ne seront pas prise en considération lors des mesures de la fécondité (D). · Cas d'une population faible: le nombre de ponte diminue, alors que le nombre d’œufs augmente par ponte (diamètre de ponte ì ). · Cas d'une population élevé: le nombre de ponte augmente, et le nombre d’œufs diminue, (diamètre de ponte î), (KHOUS, 1993). |
|
Exemple de parcelleNous avons une parcelle divisé en 03 zones selon le degrés d’attaque et importance
1 Cas d'une population faible: le nombre de ponte diminue, alors que le nombre d’œufs augmente par ponte (diamètre de ponte ì ). 2. Cas d'une population élevé: le nombre de ponte augmente, et le nombre d’œufs diminue, (diamètre de ponte î ) Le niveau de population gouverne son comportement de ponte Donc : Les masses d’œufs tendrait à augmenter de longueur avec l’abaissement du niveau de population. Les masses d’œufs deviennent plus grandes de taille et plus homogènes avec l’abaissement du niveau de population. Soit Augmentation du nombre d’œufs par masse implique diminution du nombre de ponte par arbre soit diminution de la population. Diminution du nombre d’œufs par masse implique augmentation du nombre de ponte doit augmentation de la population. |
|
ANONYME, 1973. Utilisation des virus dans la lutte contre les insectes nuisibles et vecteurs de maladies. Rapp. n°91, réunion F.A.O./O.M.S. sur les entomovirus, Rome, 41 pp. ATGEP P. & DUSAUSSOY G., 1963 Les maladies de et les possibilités de lutte microbiologique contre cet insecte: Observation sur le comportement de Calosoma sycophantha L. Rev. Pathologie Végétale et Entomologie Agricole Paris vol. XLII n°1 pp. 47-51 et 53-65. BALACHOWSKI A.S., et MESNIL L., 1935-1936 les insectes nuisibles aux plantes cultivées Paris, 2 vol. 1927 pp. BALACHOWSKI A.S., 1951 La lutte contre les insectes, principes, méthodes et application PAYOT, Paris. BARBOSA P., CAPINERA J. L., 1977 Population quality dispersal and numerical change in the gypsy moth, Lymantria dispar L. Oecologia, 36 (2) pp.: 203-209. BELABBAS D., 1991 Chêne-liège. Rev. For. Algérie n° 1, pp.: 26-30. BENAZOUN A. 1978 Bionomie d’Ooencyrtus kuwanae How; Mém. Assistanat Doc. Lab. Zool. U.A.V. Hassan II ( Rabat) 60 pp. + illust. BENMECHRI S., 1990 Présence de Brachymeria intermedia sur Tortrix viridana dans la forêt de Tamentout à Sétif. doc. interne INRF. BENCHEIKH R., 1992 Contribution à l’étude de l’écobiologie et du contrôle naturel d’Euproctis chrysorrhoea L. ( Lepidoptera, Lymantriidae ) dans la suberaie de Jijel. Mém. d’ing. d’Etat I.N.A. - Algérie 41 pp. BESS H.A., 1961 Population ecology of the gypsy moth Porthetria dispar (L.) ( Lepidoptera, Lymantriidae ). Bull. Conn. Agr. Exp. Stn., 43 pp. BOHMFALK G.T., 1986 Pratical factors influencing the utilisation of bactoviruses as pesticides. In "The Biology of Bactoviruses" Ed. Robert R. Granados and Brian A Federici. Vol. 2: pp. 223-226. CR Press, Boca. Raton, Florida (E.I.). BURGESS H.D. ET HUSSEY N.W. 1971 Microbial control of insects and mites Academic press, New-York CAMBINI A., 1972 Effecti delle defoliazioni causate dagli insetti sull’accrescimento e sull’istogenesi della quercia da sughero ( Quercus suber L.) i. Indagini anatomoistologiche sulla cerchia legnosa. Sta. Exp. Sughero. Tempio. Pausana, Mem 33: 33 pp. CAMPBELL R.W., 1974 The gypsy moth and its natural enemies U.S.Dep. Agr. Inf. Bull., 381 IV : 27 pp. CAPINERA J.L. et BARBOSA P., 1976 Dispersal of first-instar gypsy moth larvae in relation to population quality. Oecologia, 26 (1) pp.: 53-64. CHALAL N., 1993 Contribution à l’inventaire des lépidoptères défoliateurs du chêne-liège ( Quercus suber ) à Baïnem, Jijel et Djelfa. Thèse d’Ingénieur d’Etat U.S.T. de Blida, Institut d’Agronomie. CHAMBON J.P., KHOUS M.G., GENESTIER G., et PINEAU C.,1993 Contribution à l’inventaire des lépidoptères des forêts ( chênaies et cédraies ) d’Algérie Ann. For. en Algérie. COINTAT M. 1948 Remarques sur une invasion de Lymantria dispar L. Dans le Cantonnement d'UZES. Rev. For. Fr. Tome LXXXVI-Série 46e Année n°1 janv. 1948. Pp. 11-32. DAJOZ R., 1980 Ecologie des insectes forestiers. Ed. Gauthiers-Villars, Paris 489 pp. DELASSUS M. BRICHET A., BALACHOWSKY A.S., et LEPIGRE A., 1931 Les ennemis des cultures fruitières en Algérie. Moyens pratiques de les combattre. Bibliothèque du colon de l’Afrique du Nord Alger. Algérie, pp.: 139-149. DELASSUS M., 1925 La lutte contre le Liparis dispar dans le massif de l’Edough. Rev. Agric. Afr. Nord 23 : 334-336 et 348-352. DOUMANDJI-MITICHE B. & DOUMANDJI S., 1988 la lutte biologique contre les déprédateurs des cultures. Ed. O.P.U. Mars 1988. DU MERLE P., 1980 Utilisation de pièges sexuels dans une étude de la tordeuse verte du chêne, Tortrix viridana L., en montagne méditerranéenne. Les phéromones sexuelles, compte-rendu. Colmar le 25-27 novembre 1980 pp. 125-129. DUSSAUSSOY G., 1963 Observations sur le comportement de Calosoma sycophantha L. en élevage. Rev. path. Agric. de France, 42 pp.: 53-65. F.A.O./PNUD, 1986 Projet Phytosanitaire des Forêts 1986. FORBUSH E.H. et FERNALD C.H., 1986 The gypsy moth wright and potter printing co., Stade printers, Boston 49 pp. FRAVAL A. et MAZIH A., 1980 Démographie de Lymantria dispar L. Ann. Rech. For. Maroc, 20 pp.: 5-63. FRAVAL A., et HERARD F., 1975 Programme de prospection de la faune antagoniste de Lymantria dispar (L.) au Maroc U.A.V. Hassan II Maroc FRAVAL A., HERARD F. et JARRY M., 1978 Méthodes d’échantillonnages des populations de pontes de Lymantria dispar (L.) en Mamora - Maroc. Ann. Ecol. Anim. 10 (2) pp.: 267-279. FRAVAL A., GRAF P., HAMDAOUI M., KADIRI Z., RAMZI H., VILLEMANT C., 1989 « Lymantria dispar » Ed. Act. Inst. Agr. Vet. Hassen II Maroc, 219 pp. + planches. GUETTAS A. 1992 Rapport sur l’économie du liège en Algérie. doc. int. INRF. 31 pp. HAMAMI R., 1986 Note d’information sur un ravageur du chêne-liège : Lymantria dispar (L.) en Algérie Doc. int. INRF. : 8 pp. HAMRA KROUA S., 1986 Note préliminaire sur les ennemis naturels de Lymantria dispar (L.) dans les forêts de chêne-liège du Nord Constantinois. Ann. Inst. Nat. Agr. d’El-Harrach - Algérie 10 (1) pp.: 26-44. HERARD F., 1978 Comportement des adultes d’Ooencyrtus kuwanae How. ( Hymenoptera, Eoncyrtidae ), parasite oophage du Lymantria dispar L. ( Lepidoptera, Lymantriidae ), en forêt de la Mamora Ann. Zool. Ecol. Anim., 10 pp.: 603-610. HERARD F., 1979 Action des ennemis naturels de Lymantria dispar L. ( Lepidoptera, Lymantriidae ), en forêt de la Mamora. Entomophaga, 24 (2) pp.: 163-175. HERARD F., et FRAVAL A.,1980 La répartition et les ennemis naturels de Lymantria dispar L. ( Lepidoptera, Lymantriidae ), 1973-1975. Acta Oecologica, Oecol. Applicata, 1 (1) pp. 35-48. HERARD F., MERCADIER G. et ABAI M., 1979 Situation de Lymantria dispar L. ( Lepidoptera, Lymantriidae ), et son complexe parasitaire en Iran. Entomophaga, 24 (4) pp.: 371-384. HERARD F., MERCADIER G., 1980 Bionomies comparées de deux souches ( marocaine et américaine ) d’Ooencyrtus kuwanae How. ( Hymenoptera, Eoncyrtidae ), parasite oophage de Lymantria dispar L. ( Lepidoptera, Lymantriidae ). Entomophaga, 25 pp.: 129-138. JANCOVIC L., 1958 Dynamism of the gypsy moth population at Jakovacki. Kljuc. Zast. Bilja pp.: 35-46. JOBIN L., 1979 Tête à tête avec la spongieuse. Forêt Conservation 46 (1) pp.: 11-14. JOBIN L., 1982 Observations sur le développement de la spongieuse Lymantria dispar L. (Lepidoptera ) sur deux conifères de l’Ouest canadien, le douglas laxifolie et la pruche occidentale. Rev. Rech. Serv. Canadien des Forêts Vol 2. n°2 pp.: 11-13. JOBIN L., 1983 Résultats de traitements aériens à l’aide du Dimilin et du Bacillus thuringiensis pour combattre la spongieuse Lymantria dispar L. au Québec Rev. Rech. Serv. Canadien des Forêts Vol.3 n°1 pp.: 11-13. KERRIS T., 1997 Les principaux ravageurs des chênes: La spongieuse Lymantria dispar L. Communication: Journée d’étude Mai 1997 CFATS-Jijel. KERRIS Tayeb. 1988 Les Méthodes et moyens de luttes contre les Insectes nuisibles des forêts au Québec (Station de perfectionnement au Centre de Foresterie des Laurentides (CFL) Québec pp. 10-11. KHOUS M.G., 1993 Contribution à l’étude de l’écobiologie et du contrôle naturel du Lymantria dispar L. ( Lepidoptera, Lymantriidae ) en chênaie verte de Djurdjura ( Tikjda) Thèse de Magister en biologie I.S.N.; U.S.T.H.B. Algérie 133 pp. + illust. KHOUS M.G., 1993 Étude écologique des pontes du Lymantria dispar (L.) en chênaie verte de Tikjda ( Parc National ) Ann. Rech. For. en Algérie V1/93 pp.: 19-29. KHOUS M.G., VALERO J.R. et JOBIN L. 1989 La présence d’une polyédrose nucléaire chez du Lymantria dispar (L.) en Algérie Biocontrol New V.4 pp.: 54-57. KLEPAC D., 1959 Détermination de la perte d’accroissement dans la forêt attaquée par Lymantria dispar. Sumarski List. Zagreb Mazuranicev trg. br. Août - Sept. 1959. KULMAN H.M., 1971 Effects of insect defoliation on growth and mortality of trees. Ann. Revue Entomol. pp.: 289-324. LENAIL F., 1980 la lutte biologique et les organisations internationales. Phytoma, Déf. des cult. n°322 pp.: 20. LEONARD D.E., 1974 Recents developements in ecology and control of the gypsy moth. Ann. Rev. Entomol., 19 pp.: 197-229. MANOLER A 1968 A field test for the control of Lymantria dispar with noclear polyedrosis virus. Ann. Soc. Ent. Fr. ( N.S.), 4 (1) pp. 227 à 232. NENON J.P., 1981 l’utilisation des insectes entomophages en lutte biologique. ann.; Biol. T.XX, fasc.3 pp.: 228-254. PEYSSOU M., 1960 Il faut encourager la culture du chêne-liège. Revue Chêne-liège n°1582 Constantine (Algérie). Mars 1960. PODGWAITE J.D., SHIELDS K.S., SERILLO R., & BREN., 1979 Environmental persistance of the nucleopolyhedrosis virus of he gypsy moth Lymantria dispar. Eviron. Enomol., 8(3): pp. 528-536. SHAPIRO M., ROBERTSON J.L., INJAC M.J., KATAJIN J. & BELL R.A., 1984 Comparative infectivities of gypsy moth (Lepidoptera: Lymanriidae) nucleopolyhedrosis virus isolates from North America, Europa and Asia. J. Econ. Entomol., 77: pp. 153-156. SULLIVAN C.R. et WALLACE D.R., 1972 The potential northern dispersal of the gypsy moth, Porthetria dispar L. ( Lep., Lymantriidae.) Can. Entomol., 104 pp.: 1349-1355. PASTRE P. 1990 la lutte contre les ravageurs de la forêt. Dossier Delméthrine - Roussel-Uclaf. Division agrovet. pp.: 30-35 VALERO J.P., 1983 la lutte microbiologique contre les insectes nuisibles, protection des plantes en agriculture et en foresterie. Doc. int. C.F.L. Québec - Canada ZERAIA L., 1976 Quelques données sur la vie et la lutte biologique contre le Lymantria dispar CNREF, doc. Interne 4 pp. ZERAIA L., 1988 Relation entre la croissance de l’appareil aérien de Quercus suber L., et le comportement du Lymantria dispar L. Ann. Inst. Nat. Agro. Vol. 12 n° spécial pp.: 1-26. |
|
|